Пространственный контакт и порядок в ядре глазами «биолога-ядерщика»
В рамках информационного партнёрства с научно-популярным сайтом biomolecula.ru публикуем журнальный вариант статьи, которая была представлена на конкурс «био/мол/текст-2018».
До недавнего времени мы ничего не знали об ультрамикроскопической жизни генома, но с развитием науки приходим к пониманию, что эта жизнь — важный фактор управления генетическим аппаратом клетки. Теперь весь геном можно рассматривать как сеть пространственных взаимодействий генетических элементов, что принципиально изменяет наше понимание событий внутри ядра. В этой статье мы разберёмся, почему трёхмерность генома исключительно важна с точки зрения эволюции.
 Обычно мы представляем себе хромосомы в виде микроскопических Х-образных структур, плавающих в клеточном ядре. Но в таком виде хромосомы проводят лишь крошечную часть своей жизни — во время метафазы деления, и в это время никакого ядра, конечно, нет. Метафазные хромосомы содержат очень плотно упакованную ДНК, так что никакая работа с генетическим материалом невозможна, и единственная цель таких хромосом — безопасно разделить генетический материал материнской клетки пополам между двумя дочерними.
Обычно мы представляем себе хромосомы в виде микроскопических Х-образных структур, плавающих в клеточном ядре. Но в таком виде хромосомы проводят лишь крошечную часть своей жизни — во время метафазы деления, и в это время никакого ядра, конечно, нет. Метафазные хромосомы содержат очень плотно упакованную ДНК, так что никакая работа с генетическим материалом невозможна, и единственная цель таких хромосом — безопасно разделить генетический материал материнской клетки пополам между двумя дочерними.
Каждая метафазная хромосома содержит в себе две одинаковые молекулы ДНК, которые расходятся к полюсам клетки; вокруг них образуется ядерная мембрана. Внутри неё они проведут всё ближайшее время до следующего деления, и здесь они меняют конформацию — хромосомы теряют очертания, расплываются, увеличиваются в размере. Однако они не «деспирализуются» (выпрямляются), а переходят из состояния очень плотного и компактного клубка в другое, которое можно описать как всё ещё клубок, но уже неплотный, как у наушников в кармане. Только в таком виде клетка сможет работать с той генетической информацией, которая заключена в ней. Объём этой информации чрезвычайно велик: в геноме человека около 20 тысяч генов, кодирующих белки, а вдобавок ещё и гены, кодирующие разные виды служебных РНК, и, кроме того, много всякого генетического мусора, от которого по тем или иным причинам эволюция не смогла избавиться.
В каждый момент жизни клетка использует очень небольшую долю ДНК. Возникает вопрос, как клетка не теряется в массе ненужной информации и как ухитряется держать «под рукой» нужные участки генома. Заглянем в трёхмерную структуру хромосомных клубков и расшифруем пространственные контакты между разными участками ДНК. Точнее, не ДНК, а хроматина — вещества хромосом. ДНК в клетке сама по себе не живёт, с ней всегда связаны разные белки и РНК, и вместе они образуют то, что мы называем хроматином.
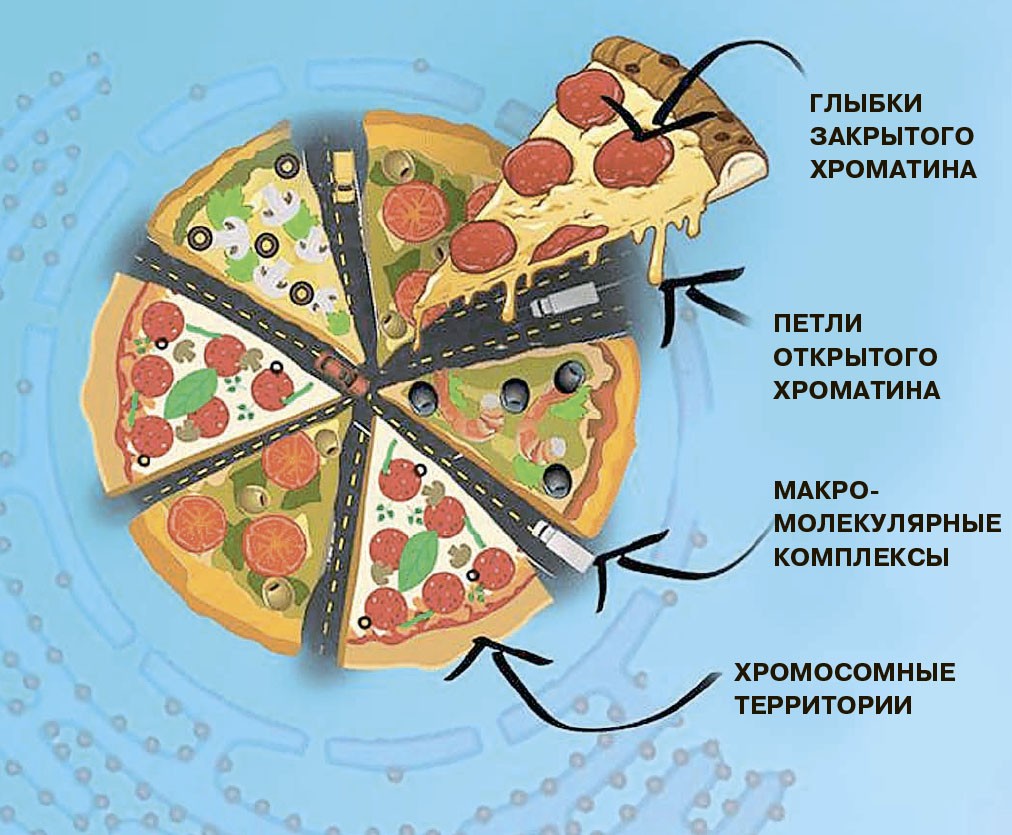
В очень маленьком объёме ядра собрано огромное количество молекул. Формируются макромолекулярные скопления, когда крупные молекулы собираются в агрегаты под действием энтропийных сил, то есть без затрат энергии. На пальцах это можно объяснить так: в жидкости маленькие молекулы двигаются по-броуновски хаотично и при соударении с крупными молекулами передают им импульс. Если рядом с макромолекулой нет других макромолекул, то она испытывает давление со всех сторон и остаётся в покое. Если же к ней случайно приблизится другая макромолекула, то со стороны, где они сближаются, небольшие молекулы будут реже их толкать; следовательно, уменьшится давление на макромолекулы — и они сблизятся ещё сильнее. Так получаются макромолекулярные скопления в условиях «перенаселённого» клеточного ядра.
В таких скоплениях разные участки хроматина могут оказываться далеко друг от друга, а могут — и рядом друг с другом. Случайным ли будет такое соседство? Совсем нет. Дело не только в густоте молекулярного «супа» и энтропийных силах. Белки хроматина, которые играют в его жизни главную роль, ведут себя очень активно: они садятся на ДНК, ползают по ней, таскают её в разных направлениях, склеиваются друг с другом и отклеиваются. В их поведении нет даже намёка на случайность — они точно контролируют положение ДНК в ядре, и от них зависит вероятность пространственного контакта между любыми двумя хроматиновыми зонами. Чтобы оценить эту вероятность, есть специальные методы — методы захвата конформации хромосом, например метод Hi-C. С помощью таких технологий можно увидеть, что пространственная структура хроматина в ядре повторяется у разных клеток. Значит, у пространственных контактов между разными участками хроматина есть функциональный смысл.
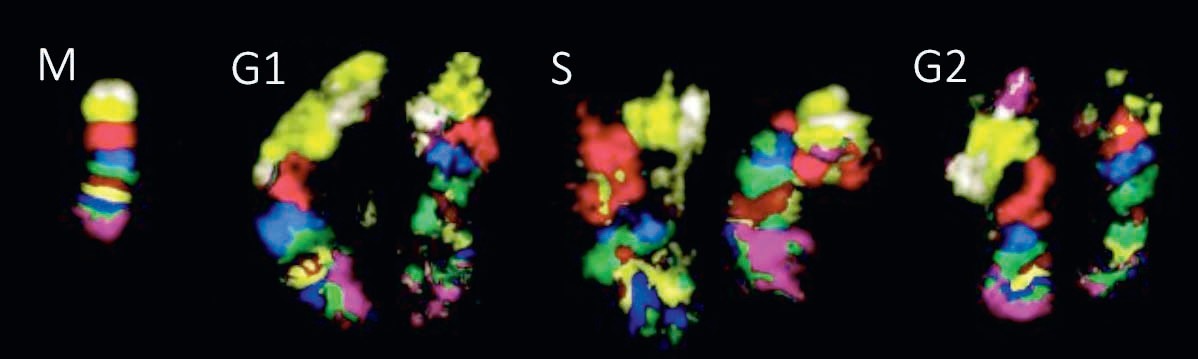
Какие тут закономерности? Во-первых, когда хромосомы после деления из конденсированной, плотно упакованной палочки превращаются в расплывчатый клубок (деконденсируются), они не перемешиваются друг с другом и не распределяются равномерно по всему ядру. Методами микроскопии и методом Hi-C удалось показать, что хромосомы занимают ограниченные и к тому же неперекрывающиеся области, получившие название «хромосомные территории». То есть хромосомы не перемешиваются друг с другом и внутрихромосомные контакты намного более вероятны, чем межхромосомные. Даже гомологичные хромосомы — то есть те, которые несут одинаковые гены, но достались от разных родителей, — в человеческих клетках занимают разные участки, которые могут и не соседствовать. И если внутрихромосомные контакты обычно воспроизводятся после деления, то межхромосомные — как правило, нет, так как хромосомные территории распределяются в каждом клеточном цикле случайным образом.
Во-вторых, если мы рассмотрим уже внутрихромосомные контакты, то обнаружим, что активные участки ДНК контактируют с такими же активными участками и избегают контактов с неактивными и наоборот. Что значит «активные участки»? Это те, которые доступны для транскрипции, то есть для считывания генетической информации, для копирования её в молекулы РНК. Следовательно, клетка пространственно отделяет используемые и неиспользуемые участки генома. Ключевую роль здесь играют белки-гистоны, которые служат упаковщиками ДНК. Гистоны слипаются друг с другом, но, как именно они будут слипаться, зависит от их химических модификаций. Модификации могут быть разные; одни из самых распространённых — это метильные или ацетильные группы, прикреплённые к определённым аминокислотам в гистоновой молекуле. Нужная генетическая информация отделяется от ненужной с помощью метилирования или ацетилирования гистонов. В общем случае вместе слипаются белковые молекулы, помеченные метильными группами и не помеченные ничем, а вот ацетилированные гистоны, наоборот, пытаются отделиться от метилированных. Гистоны с метильными группами сидят на тех участках ДНК, где хранится ненужная информация, и этот хроматин, грубо говоря, «выпадает в осадок» — получаются так называемые хроматиновые глыбки. ДНК, содержащая активные гены и регуляторные элементы для них, «всплывает» над неактивным хроматиновым «осадком».
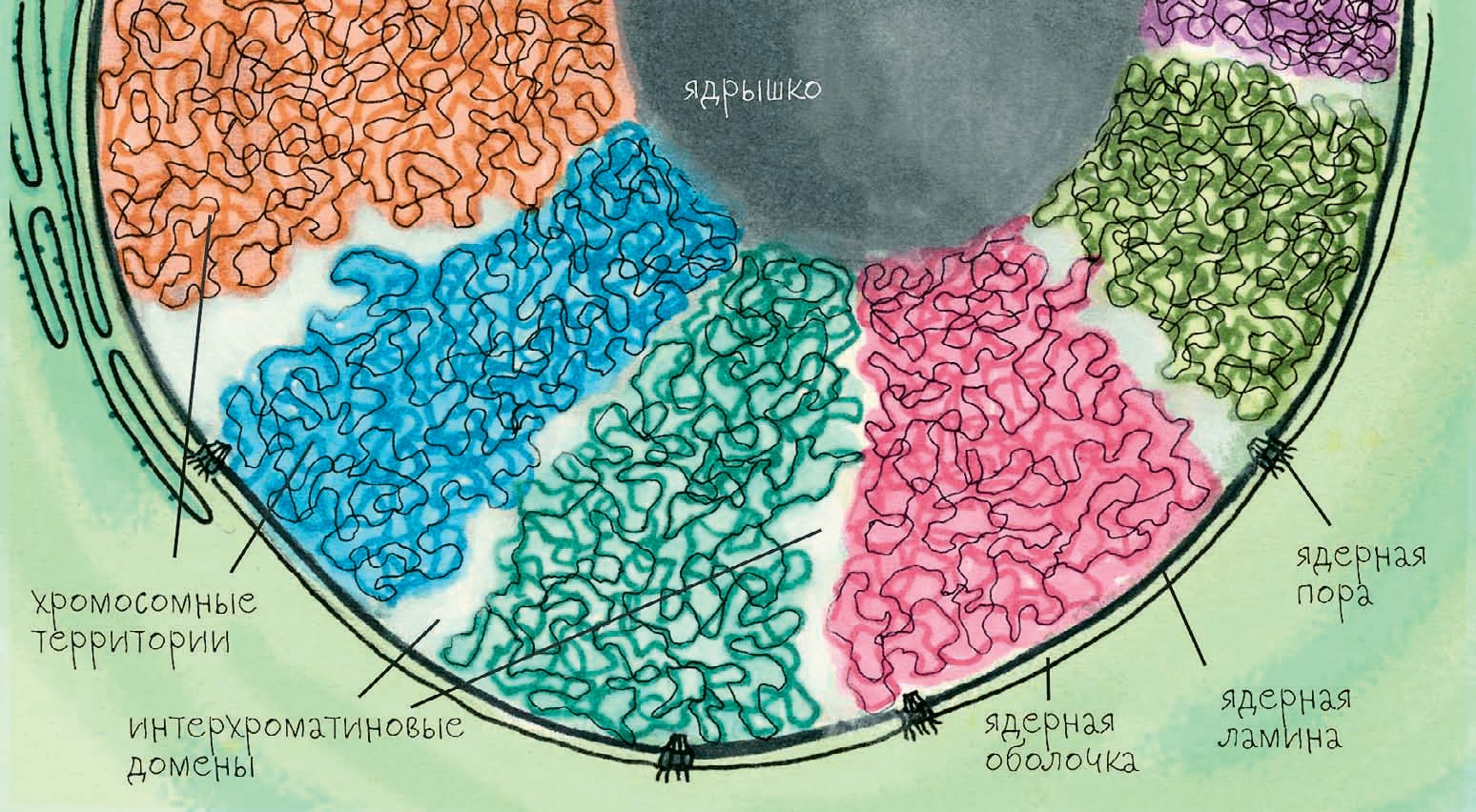
Хроматиновые глыбки окружены интерхроматиновым компартментом — ядерным матриксом с функциональными агрегатами, большими молекулярными структурами, которые заняты синтезом РНК и её последующей обработкой — редактированием. Интерхроматиновое пространство можно рассматривать как сеть тоннелей, начинающихся от открывающихся в цитоплазму ядерных пор и пронизывающих всё ядро. Крупные белковые комплексы не могут проникнуть внутрь хроматиновых глыбок, так что вся ядерная молекулярная работа происходит именно в этих интерхроматиновых тоннелях. В тоннели же обращены ацетилированные (то есть активные) участки хроматина — они покрывают глыбки «ворсом» из петель ДНК. Ненужная информация контактирует сама с собой внутри плотно упакованных глыбок, а нужные фрагменты генома выведены из глыбок наружу, в интерхроматин, где они могут контактировать друг с другом и с белками, обслуживающими нужную генетическую информацию. Таким образом клетке удаётся без особых затрат времени и сил поддерживать свой геном в порядке даже после деления.